21
Conclusions
Lacompréhensiondu systèmeolfactif des insecteset de
saplasticitépermet ledéveloppementd’applicationsdans
plusieursdomaines.Enagronomie, lespiègesolfactifspour
lutter contre les ravageursdescultures sont déjàutilisés ;
ensantéhumainedesméthodesdeperturbationdesvec-
teursdemaladiesdans leur recherched’hôtesousitesde
reproductionsont enpleinessoret, enolfactionartificielle,
des solutions biomimétiques des stratégies d’orientation
des insectessont testéessurdes robots traqueursd’odeur.
B
ibliographie
(1) Martin J.P., et al. (2011) ProgNeurobiol 95(3):427-447.
(2) Keil T.A., in Insect olfaction, B.S.Hansson, Editor. 1999, Springer:
Berlin. p. 5-47.
(3) LealW.S. (2013)AnnuRevEntomol 58:373-391.
(4) KauppU.B. (2010)Nat RevNeurosci 11(3):188-200.
(5) GaliziaC.G. et RosslerW. (2010)AnnuRevEntomol 55:399-420.
(6) Schachtner J., SchmidtM., etHombergU. (2005)ArthropodStruct
Dev34(3): 257-299.
(7) MasseN.Y., TurnerG.C., and JefferisG.S. (2009)CurrBiol
19(16):R700-R713.
(8) Jeanne J.M. etWilsonR.I. (2015)Neuron88(5):1014-1026.
(9) BustoG.U.,Cervantes-Sandoval I., etDavisR.L. (2010) Physiology
(Bethesda) 25(6):338-346.
(10) GadenneC., BarrozoR., et AntonS. (2016)AnnuRevEntomol
61:317-333.
L’olfaction chez les amphibiens
Jean Gascuel
(Centredes Sciences duGoût et de
l’Alimentation, CNRSUMR6265, Dijon)
Chez laplupartdesespèces,depuis les insectes jusqu’aux
mammifères, l’organisation générale du système olfactif
présentedenombreusessimilarités
(1,2)
.Cesdernièressug-
gèrent que l’organisationdusystèmeolfactif est largement
conservée au cours de laphylogenèse. En conséquence,
enolfactioncommedansd’autresdomaines, laplupartdes
recherches sont actuellement concentrées sur un nombre
très réduit demodèles : la souris, lepoisson zèbre, ladro-
sophileet le vers
Caenorhabditis
.
Sans douter de l’utilité et d’une forme de représentativité
decesmodèles, il s’avèrequ’àcôtédes traitsgénéraux -et
largement partagés - qui caractérisent l’organisation et la
physiologiedessystèmesolfactifs, une incroyablediversité
estévidente lorsqu’onanalyseunpaneld’espècesplus large
(3)
. Si l’on acceptedeprendre en compte cettediversité,
certainsmodèlesoffrentdesparticularitésd’organisationou
dephysiologie toutà faitétonnantes,quipermettentdeposer
uncertainnombred’hypothèses fonctionnellesdemanière
trèspertinente.C’est justement lecasdesamphibiens.
Chez les amphibiens, l’olfaction intervient dans tous les
comportements tels que la reproduction, la recherchede
nourriture, lecomportementd’agrégation, lemarquage ter-
ritorial. La physiologie de la détection olfactive au niveau
périphérique et central a largement été étudiée chez la
grenouille
(4)
et chez lexénope
(pour revue, 5)
.Cedernier
modèleoffre lacaractéristique, sansdouteuniquedans le
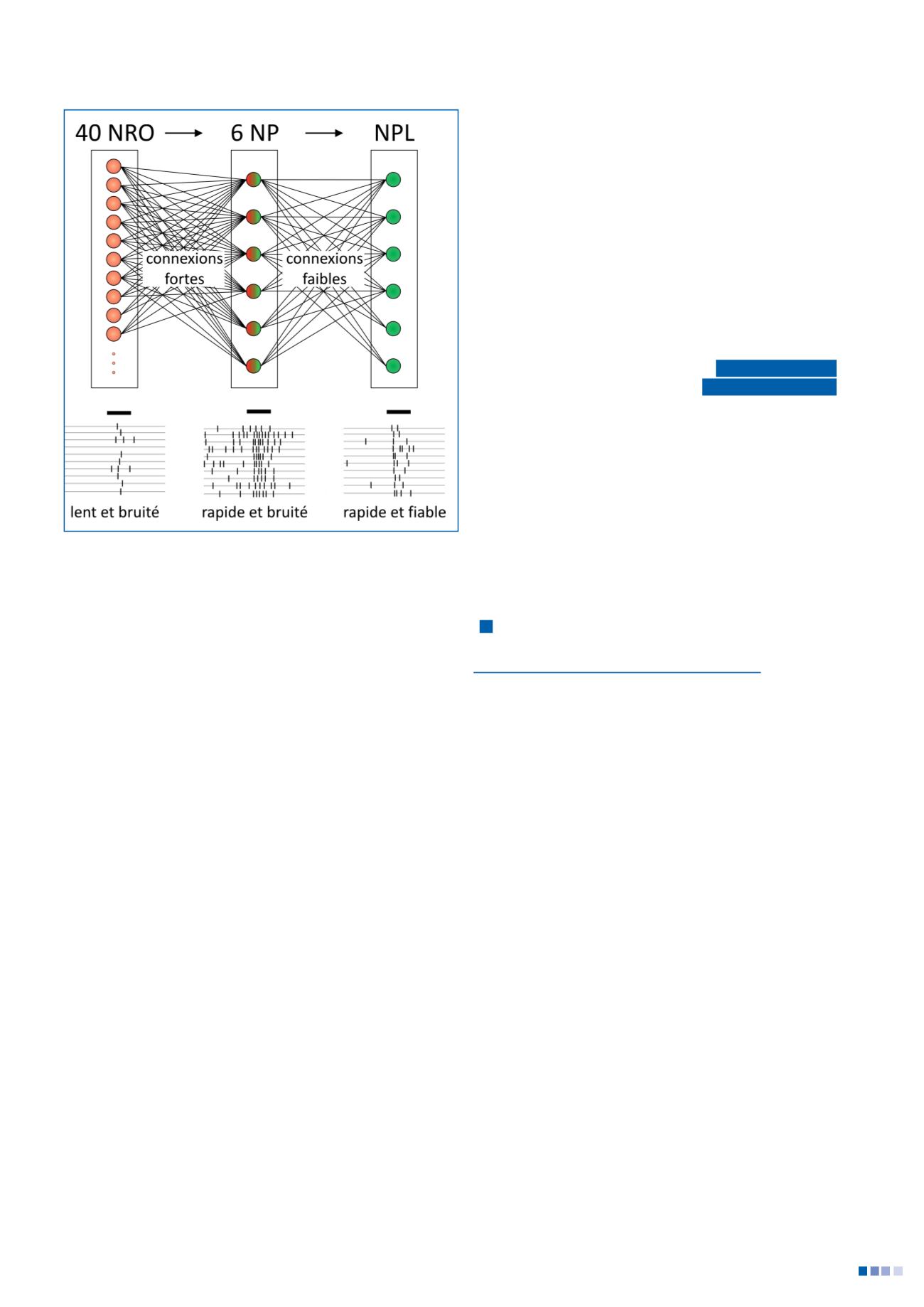
sont trèssynchronisées, tout comme les réponsesdesNPL.
LecouplagesynaptiqueNP-NPLest faibleet lesNPLont un
seuil d’émissiondepotentielsd’actionélevéet dynamique.
LesNPLsedépolarisentplusvitequand l’activitédesNPest
synchronisée (lorsd’une réponse)quedésynchronisée (acti-
vitéspontanée). Il en résulteque les réponsesdesNPLsont
rapideset fiablesen raisonde leuractivitéspontanée faible.
Ainsi, les signaux bruités et asynchrones délivrés par les
NRO sont intégrés pour produire un signal sortant dupro-
tocérébron latéral qui est à la fois rapideet fiable.
Plasticitédusystèmeolfactif
Lesystèmeolfactif des insectes,bienquemoinscomplexe
queceluidesvertébrés,gardeunegrandeflexibilitéetadap-
tabilitéà leurenvironnement changeant.Chezdes insectes
modèles, comme l’abeille, lespapillonsdenuit ou ladroso-
phile,différentes formesdeplasticitédusystèmeolfactif ont
étémisesenévidence.L’étatphysiologiquede l’insecte (âge,
statut viergeouaccouplé)maisaussi l’expérienceolfactive
(habituation, sensibilisationouapprentissagepar associa-
tion)changent les réponsescomportementalesdes insectes
auxsignauxolfactifs.Cesmodifications induisent,par l’inter-
médiaired’hormonesetaminesbiogènes,deschangements
anatomiques et physiologiques dans les centres olfactifs
cérébrauxprimaireset secondaires, et plus rarement aussi
au niveaudesNRO
(9,10)
. Lesmécanismes cellulaires et
moléculaires de ces changements sont étudiés endétail,
surtout chez l’abeilleet ladrosophile.
Figure2
-Circuits impliquésdans lesréponsesaucVAchez la
drosophile.Les signauxbruitésetasynchronesdélivréspar les
neuronesrécepteursolfactifs (NRO) sont intégréspar lesneuronesde
projection (NP), pourproduireun signal sortantduprotocérébron
latéral (NPL)qui està la foisrapideetfiable (d’après (8)).


