9
Quelques réflexions d’un vieux physio-
logiste
Michel Jouvet
(membre de l’Institut)
Mon ami Yves Tillet pratique une gérontologie offensive en
demandant au presque nonagénaire que je suis (né en 1925)
de donner son avis sur l’évolution actuelle de la neurobiologie
du rythme éveil, rêve, sommeil.
Qu’il en soit remercié, car la lecture du Dossier de La Lettre
de la Société des Neurosciences m’a vivement intéressé et
suggéré les réflexions suivantes :
1) Il me semble que l’école lyonnaise continue à se situer
au tout premier rang de la recherche sur l’exploration des
mystères du cycle Éveil-Rêve-Sommeil.
2) Les réseaux neuronaux (voir l’article de P.H. Luppi) sont
maintenant bien connus (en attendant la découverte de
nouveaux transmetteurs) et le physiologiste que je suis
se demande pourquoi, au cours de l’évolution, tant de
systèmes et de neurotransmetteurs se sont intriqués ? Il est
vrai que l’éveil est le comportement vital par excellence.
Il existe des animaux ou des malades qui peuvent ne pas
dormir pendant des mois (voir plus loin) mais on ne connaît
pas d’animaux qui dorment tout le temps.
Bien sûr, la pharmacologie a déjà permis d’agir sur l’acti-
vité des systèmes monoaminergiques avec les inhibiteurs
des monoamines oxydases et de supprimer ainsi le som-
meil paradoxal (SP) sans altérer l’éveil. Les souris sans
histamine, de J.S. Lin et son équipe, semblent incapables
de rester éveillées dans un nouvel environnement. Cela
leur est possible dans un laboratoire, mais les condam-
nerait à la mort si elles étaient en liberté.
Mais l’exploration de la phyloge-
nèse des réseaux de l’éveil, de-
puis les mollusques, nématodes,
annélides, jusqu’aux poissons,
crocodiles, oiseaux et de l’onto-
genèse depuis les fœtus de rats
ou de chats, permettrait sans
doute de comprendre pourquoi
l’évolution a inventé un tel réseau.
3)
Sommeil et consolidation mné-
sique
(voir l’article de G. Rauchs
de Caen).
Année après année, on voit ap-
paraître des publications pour
(soutenues surtout par l’Ecole de
Harvard) et contre le rôle géné-
tique du sommeil dans la consoli-
dation mnésique. Les travaux des
auteurs de Caen sont originaux
et s’appuient sur l’activité hippo-
campique. J’ai ainsi appris qu’au
cours du sommeil mon cerveau
serait capable de discriminer des
odeurs, des sons et même un
prénom. Mais à quelle intensité
sonore ?
Sommeil et mémoire, j’aimerais y croire mais je vous invite
à lire la thèse du Dr Fisher-Perroudon (1974), qui résume
une observation onirique : un malade de 23 ans atteint
d’une « agrypnie » (absence totale de sommeil)
pendant
4 mois
dont le sommeil était enregistré de façon perma-
nente lorsqu’il se couchait avec un appareil EEG. Malgré
cette totale absence de sommeil (sommeil léger, stade
II, III et IV et SP), ce malade n’a présenté
aucun trouble
de l’attention ni de la mémoire
(apprécié par des tests
pluri-mensuels).
Si bien que la conclusion de cette étude fut la suivante :
A quoi sert le sommeil ?
4) Je retiendrai de l’article de mon ami André Malan, l’extra-
ordinaire référence publiée en 2000. La réanimation sans
séquelle neurologique d’une hypothermie accidentelle à
13,7°C chez une skieuse après un arrêt circulatoire de
plus de 2 heures.
Et pourquoi ne me ferais-je pas hiberner pendant quelques
années (sous contrôle par des spécialistes) pour pouvoir
assister, dans les années 2025, à la solution possible des
mystères du sommeil et du rêve ?
Les réseaux neuronaux du cycle
veille-sommeil
Pierre-Hervé Luppi
(Univ. Claude Bernard Lyon 1)
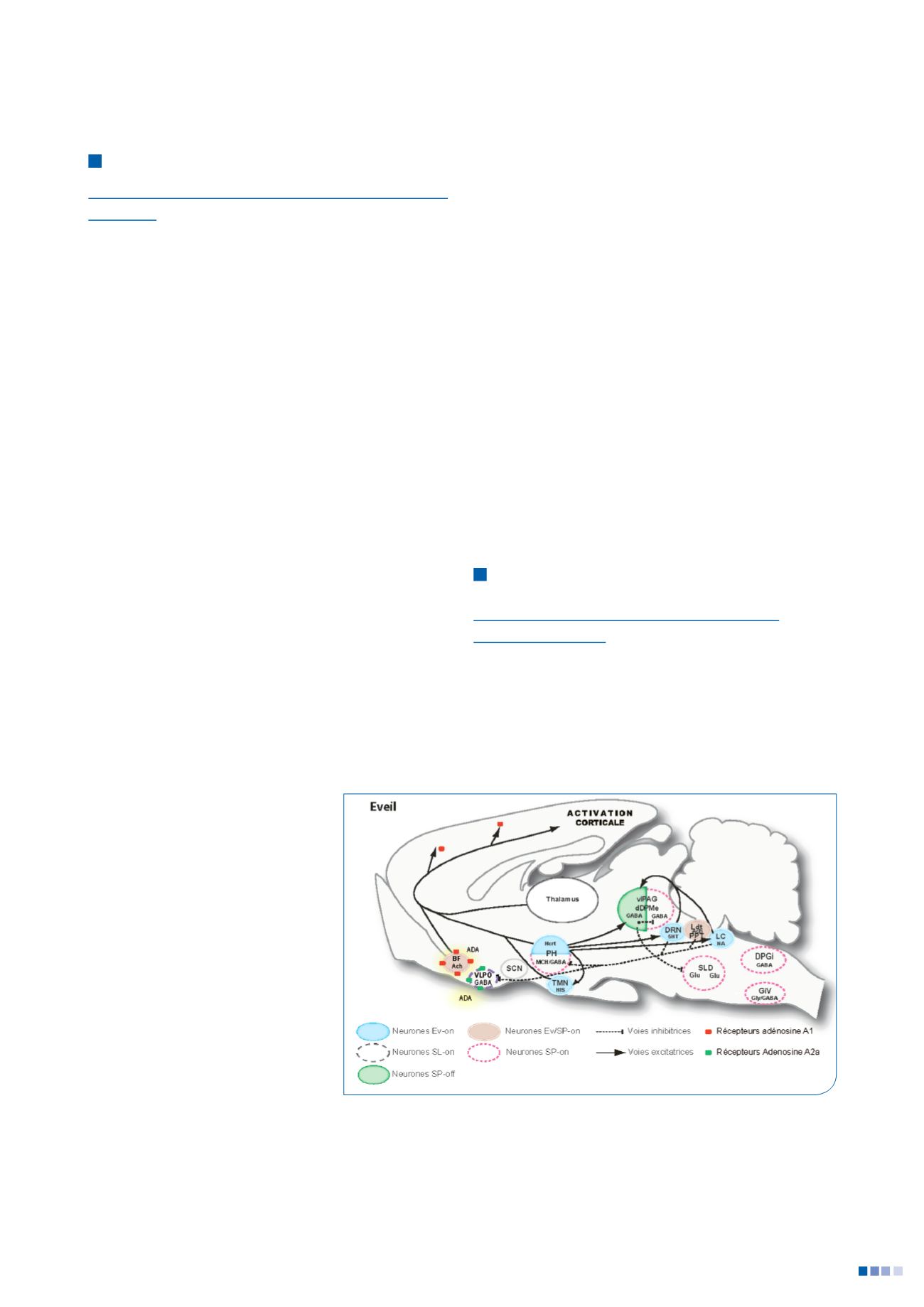
1. Réseau de l’éveil
L’éveil est caractérisé par une activation corticale et une
activité motrice tonique et phasique. Les études de lésions
Figure 1 :
schéma du réseau de l’éveil
Voir détails dans le texte.
Abréviations : 5HT, sérotonine; Ach, acétylcholine; BF, Télencéphale basal, DPGi, noyau
réticulé dorsal paragigantocellulaire; dDpMe, Noyau profond réticulé mécencephalique
dorsal; DRN, noyau raphé dorsal; GiV, noyau réticulé ventral gigantocellulaire; Glu,
glutamate; Gly, glycine; Hcrt, hypocretin- (orexin); His, histamine; LC, locus coeruleus;
Ldt, noyau tegmental laterodorsal ; LPGi, noyau lateral paragigantocellulaire;
MCH, hormone de mélanoconcentration; PH, hypothalamus posterieur; PPT, noyau
pedunculopontin; vlPAG, substrance grise périaqueducale ventrolatérale; VLPO, noyau
ventrolatéral préoptique; SCN, noyau suprachiasmatique; SLD, noyau sublatérodorsal.


