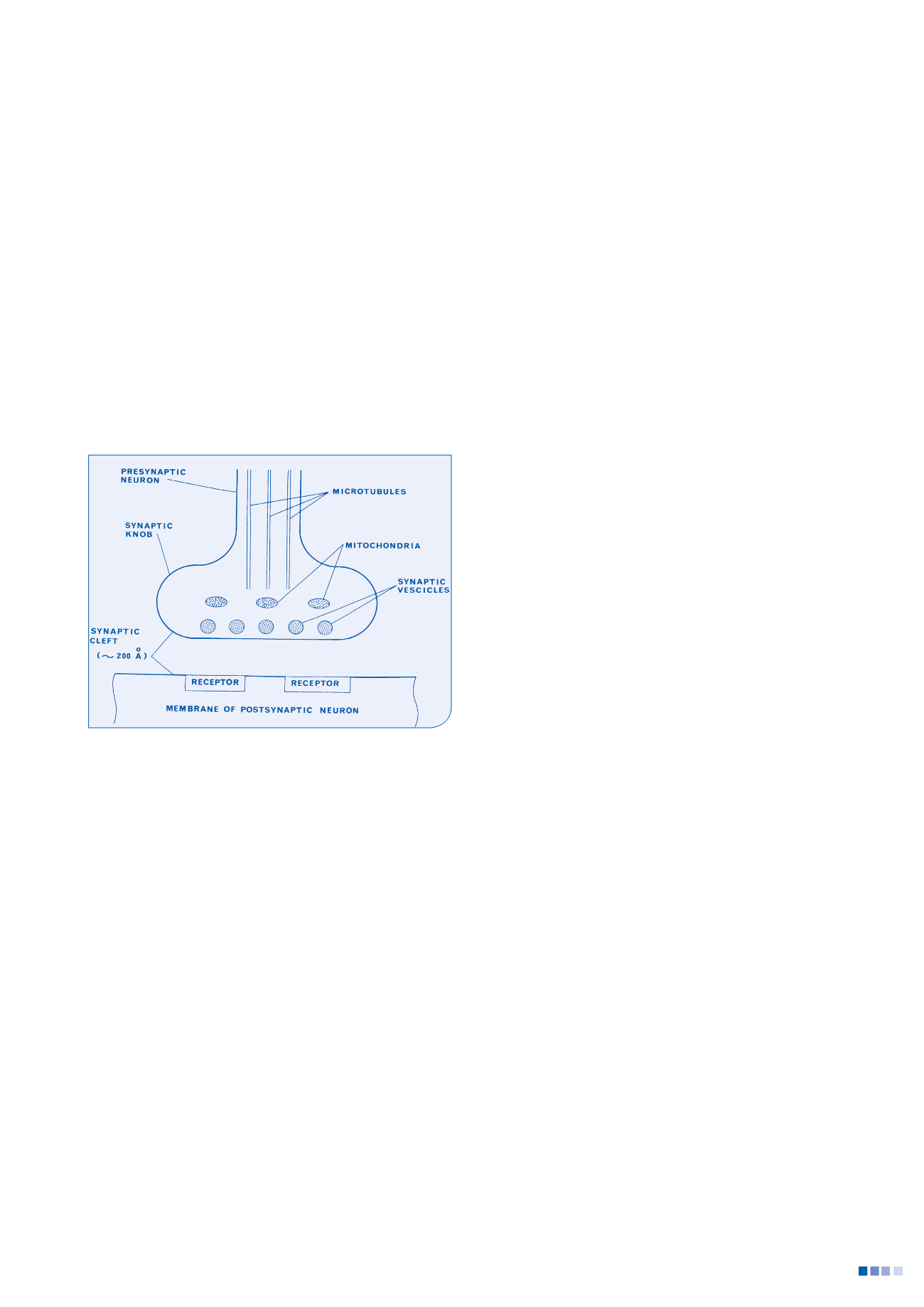
5
La vision de la synapse et de la place d’un récepteur dans la neurotransmission
chimique des années soixante-dix (The Principles of Physiology D. Jensen,
Appleton-Century-crofts, 1976)
teurs de l’action de l’AMPc. Au début des années 70, Rodbell
propose l’existence d’une protéine de couplage GTP-dé-
pendante ou protéine G servant comme traducteur entre les
récepteurs au glucagon et l’adénylyl cyclase. Rodbell décrit
un modèle explicatif de complexe ternaire ligand, récepteur
et protéine G, guidé par les constantes de dissociation à
l’équilibre entre chacun des partenaires et un facteur de
coopérativité
a
. Ce modèle suppose un état de haute affinité
du récepteur (couplé à la protéine G) et de faible affinité (non
couplé) très largement utilisé aujourd’hui, malgré ses limites.
Les années 80 : la caractérisation biochimique des
récepteurs, le clonage.
En plus des techniques de liaison, le développement d’autres
méthodologies biochimiques a contribué largement à passer
du concept de récepteur à sa description biologique : tech-
niques de marquage d’affinité, solubilisation des protéines
membranaires par des détergents, chromatographie d’affinité
pour la purification et enfin reconstitution des protéines dans
des vésicules lipidiques. Il existe très peu de sources enri-
chies en récepteurs ; plusieurs modèles d’étude s’imposent
à cette époque ; le récepteur ß2 adrénergique, le récepteur
nicotinique et la rhodopsine ; les deux derniers parce qu’on
peut obtenir des préparations presque pures à partir de
l’organe électrique de la torpille et de bâtonnets rétiniens,
respectivement.
La capacité de marquer directement des sites de liaison a
aidé grandement à la purification biochimique de récepteurs.
Le groupe de Lefkowitz en 1988
(4)
purifie l’adrénorécepteur
ß2 de hamster ; la séquence partielle d’amino-acides obtenus
par clivage au CNBr permet de cloner le gène. Il est composé
d’une chaîne polypeptidique unique, plus ou moins glyco-
sylée et phosphorylée, avec une masse de 60 000 daltons.
Les ß-bloquants se lient à la protéine réceptrice isolée avec
la spécificité appropriée et de manière stéréospécifique. Le
gène codant pour le récepteur ß2-adrénergique est sans
intron ; plus surprenant, il contient sept segments de résidus
hydrophobes similaires aux segments transmembranaires
proposés pour la rhodopsine. Cette dernière, séquencée
en 1982 par les méthodes biochimiques conventionnelles,
possède sept hélices transmembranaires et se lie à une
protéine G de manière lumière dépendante.
De 1982 à 1983, Shosaku Numa au Japon, publie trois articles
sur l’identification de quatre gènes codant pour les différentes
sous-unités du récepteur nicotinique de l’électroplaque de
la torpille électrique. Une séquence de 55 amino-acides,
précédemment identifiée par Raftery, a été suffisante pour
construire une séquence d’oligonucléotides et l’identification
de quatre gènes distincts, qui, exprimés dans des ovocytes
de xénope confirment l’existence d’un récepteur canal entier
fonctionnel. L‘utilisation de sondes oligonucléotidiques pour
« pêcher » les ADN complémentaires codant pour des récep-
teurs devient la technique de la décennie.
On étudie les propriétés de liaison de tous ces nouveaux
récepteurs pour expliquer la diversité des réponses biolo-
giques ; on révèle des sous-types pour tous ces récepteurs,
les processus de régulation négative, les modifications post-
Les années 70 sont, pour les États-Unis, celles de la guerre au
Vietnam et la peur d’une épidémie d’usage abusif d’héroïne
par les vétérans ; on étudie donc les opiacés qui agissent à
très faibles doses, de manière stéréospécifique et qui pos-
sèdent probablement des sites de liaison. Un antagoniste
tritié avec une très haute activité spécifique : la naloxone,
un système de filtration sur filtre de verre pour séparer le
ligand lié du libre, un lavage pour minimiser la quantité de
liaison non-spécifique et les récepteurs spécifiques des
opiacés sont identifiés en 1973, parmi les tout premiers. On
les localise dans des aires du cerveau impliquées dans la
perception douloureuse, l’euphorie, la dépression respira-
toire ; tous les aspects des activités pharmacologiques de
la morphine dont on ignorait jusqu’à présent la cible, sont
soudain compréhensibles.
Les sites de liaison des morphiniques caractérisés par plu-
sieurs équipes entre 1971 et 1973 (E. Simon, S. Snyder et
L. Terenius), ont permis rapidement l’isolement de subs-
tances endogènes analgésiques morphinomimétiques (les
enképhalines) en purifiant des extraits de cerveau par chro-
matographie. L’ère des neuropeptides s’ouvre. De quelques
neuropeptides connus en 1970, on passera à plus de cin-
quante, une décennie plus tard.
La même stratégie a été appliquée à beaucoup d’autres
drogues actives sur le système nerveux et a conduit à l’identi-
fication de la plupart des récepteurs aux neurotransmetteurs.
Nous voilà donc bien vite avec des cibles enfin repérables et
localisables. Ces sites de liaison identifient des cibles mais
pas encore un mécanisme. Comme toujours, des éléments
disparates s’assemblent pour former un schéma général
pour ce qu’on va appeler les récepteurs couplés aux pro-
téines G (RCPG).
Dans les années 60, Sutherland propose l’AMPcyclique
comme second messager dans l’action de plusieurs ré-
cepteurs ; les protéines kinases AMPc-dépendantes (PKA)
découvertes en 1968 complètent le tableau comme effec-


